|
FATTY ACID SYNTHESIS |
·
Not an exact reversal of β-oxidation
reactions.
·
Occurs in cytosol.
·
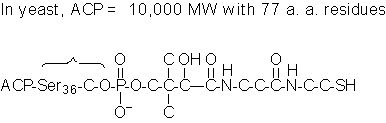
Using
an Acyl Carrier Protein (ACP or ACP-SH) instead of CoA for
the activation of FAs.
·
CO2 (as HCO3-) is required but
is not incorporated into the elongated FA product.
·
Sequential addition of 2-C units derived from acetyl-CoA.
·
The activated unit for the 2-C addition is a 3-C unit in the
form of malonyl-ACP.
·
Citrate is required for
maximal activity.
·
NADPH is required as a
reductant.
·
Elongations stop at palmitate
(C16). Further elongation and/or
insertion of double bonds require additional enzymes.
SOURCES OF ACETYL-CoA
·
Cytoplasmic acetyl-CoA synthetase
CH3-COO- +
ATP + CoA → CH3-CO-CoA + AMP
+ PPi
·
Mitochondrial pyruvate
dehydrogenase complex
Pyruvate + NAD+ + CoA → Acetyl-CoA + NADH + CO2 + H+
·
Mitochondrial b-oxidation of FAs
·
Mitochondrial degradation of some amino acids
TRANSPORT OF ACETYL-CoA (Fig. 19-24): Tricarboxylate transport system
Note: Malate can also be transported across inner
mitochondrial membrane. But this latter
shuttle will not generate NADPH in cytosol (also see discussion below).
SOURCES OF NADPH
1. Tricarboxylate transport system
·
Cytosolic malate
dehydrogenase:
oxaloacetate +
NADH + H+ → malate + NAD+
·
Cytosolic malic enzyme: malate + NADP+ → pyruvate + CO2 + NADPH
·
pyruvate
is transported into mitochondria
·
Mitochondrial pyruvate
carboxylase:
pyruvate
+ CO2
+ ATP + H2O → OAA + ADP + Pi + 2 H+
·
NADP+ + NADH + ATP
+ H2O → NADPH + NAD+ + ADP + Pi + 2 H+
·
For every cycle of the tricarboxylate
transport system, one acetyl-CoA and one NADPH are
gained in cytosol. At the same time,
one NADH plus one ATP are consumed in cytosol and one ATP plus one acetyl-CoA are consumed in mitochondrion.
2. Pentose phosphate
pathway.
Role of Citrate
1. Activator
for acetyl-CoA carboxylase
(see below) - THE committed step and
A rate-limiting Enz
for FA synthesis.
2. Citrate
functions in the transport of acetyl-CoA from
mitochondria to cytosol - the Citrate Shuttle (same as that shown in Tricarboxylate Transport System).
=============================================================
Acetyl-CoA carboxylase (in cytosol)
Acetyl-CoA + H*CO3- + ATP → -OO*C-C-CO-CoA + H+ + ADP + Pi
malonyl-CoA
·
Allosteric:
Requires citrate
or isocitrate for activation. Inhibited
by palmitoyl-CoA.
·
Requires
CO2.
(or bicarbonate HCO3-)
·
The committed step (= the first irreversible step in a pathway) and one of the rate-limiting
steps.
Fatty Acid Synthase
·
In
higher organisms, as a single polypeptide chain containing 7 activity domains.
·
In
bacteria, the constituent enzymes of fatty acid synthase are dissociated upon
isolation.
One Cycle of Elongation (See Fig. 19-26)
1a. Malonyl-CoA/Acetyl-CoA-ACP transacylase
Acetyl-CoA + ACP ⇌ Acetyl-ACP + CoA
1b. Malonyl-CoA/Acetyl-CoA-ACP transacylase
Malonyl-CoA + ACP ⇌ Malonyl-ACP + CoA
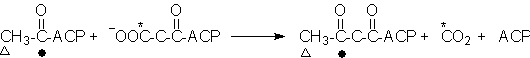
2a+2b. β-Ketoacyl-ACP synthase
(Condensing enzyme)
3. β-Ketoacyl-ACP reductase
·
NADPH-dependent, contrary to β-oxidation.
·
Produces
D-β-hydroxyacyl-ACP (or 3-D-hydroxyacyl-ACP), contrary to L-β-hydroxyacyl
group in β-oxidation.
4. β-Hydroxyacyl-ACP dehydrase
·
Produces Δ2-trans-enoyl-ACP.
5. Enoyl-ACP
reductase (→ Butyryl-ACP → ´ 6 for Palmitoyl-ACP)
·
NADPH-dependent, contrary to β-oxidation.
Recyle 6 more times to obtain Palmitoyl-ACP
6. Palmitoyl
thioesterase
Palmitoyl-ACP + H2O à R-COO- + ACP
|
See Fig. 19-23 for comparison of FA synthesis versus
oxidation. |
ENERGETIC CONSIDERATION
OXIDATION OF PALMITATE
130 ATP are formed
from the complete oxidation of palmitate (see earlier
notes).
SYNTHESIS OF PALMITATE
·
7 Acetyl-CoA + 7 CO2 + 7 ATP → 7 Malonyl-CoA + 7 ADP + 7 Pi + 7
H+
·
Acetyl-CoA + 7 Malonyl-CoA + 14
NADPH + 7 H+ → Palmitate + 7 CO2 + 14 NADP+ + 8 CoA + 6 H2O
·
8 Acetyl-CoA + 7 ATP + 14 NADPH →
Palmitate
+ 14 NADP+ + 8 CoA + 6 H2O + 7 ADP + 7 Pi
8 Acetyl-CoA → into
mitochondria → TCA → 8 ´ 12 = 96
ATP
7 ATP
7 ATP
14 NADPH → 14 NADH → Glycerol 3-P
shuttle → 14 FADH2 → 28 ATP
or → Malate-Aspartate shuttle → 14 NADH → 42 ATP
Total
= 131 or 145 ATP
FURTHER ELONGATION OF PALMITATE
1. Mitochondria
See Fig. 19-29 -
Last step requires NADPH instead of FADH2, otherwise identical to reversal of FA oxidation.
2. Endoplastic
Reticulum
Acyl-CoA (Cn) + Malonyl-CoA
→ Acyl-CoA (Cn+2) → → Similar to normal
FA synthesis except uses CoA instead of ACP.
UNSATURATION
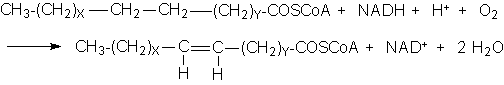
Desaturases
·
X: At least 5C, could contain double bonds.
·
Y: Always saturated.
·
e.g. Stearoyl-CoA (C18) → Oleoyl-CoA (cis-Δ9)
·
Oleoyl-CoA can be further
elongated and unsaturated to obtain poly-unsaturated FAs.
·
Mammals have Δ9-, Δ6-, Δ5-, and Δ4-fatty acyl-CoA
desaturases but cannot generate double bonds beyond
C-9.
·
Therefore, FAs such as linoleate
(18:2, cis-Δ9,Δ12) and linolenate (18:3, cis-Δ9,Δ12,Δ15) are ESSENTIAL FAs for mammals.