PHOTOSYNTHESIS |
·
Photosynthesis: The light-driven synthesis of carbohydrate (CH2O)
from CO2.
·
Photosynthetic
systems also produce O2 (from oxidation of water) for the earth atmosphere.
light
CO2 + 2 H218O ® CH2O + 18O2 + H2O
CHLOROPLASTS (Fig. 18-1)
·
The site of
photosynthesis in eukaryotes (algae and plants).
·
~1-1000
chloroplasts per cell, variable in size and shape, but typically as ~5-mm long ellipsoids.
·
Contain 3
distinct types of membranes, namely outer, inner, and thylakoid
membranes. The light-driven reactions take place in the thylakoid membrane.
·
Three separate
spaces: intermembrane
space, stroma, and thylakoid
lumen. The dark reactions take place in
the stroma.
·
Thylakoid membranes arise from invaginations of the inner membrane of
developing chloroplasts, and resemble mitochondrial cristae.
·
The thylakoid vesicle is arranged to have disk-like sacs,
referred to as grana, stacked in piles, which are
connected to each other by stroma lamellae.
·
A chloroplast
usually contains 10-100 grana.
·
Stroma, a concentrated solution containing enzymes, DNA, RNA, and ribosomes.
CHLOROPHYLLS
·
Magnesium
porphyrins.
·
Different
chlorophylls have different absorption spectra.
|
Two Phases of Photosynthesis |
1. Light Reactions
·
Use light energy to generate NADPH, ATP and O2.
·
Occur in the thylakoid membrane, involving processes resembling ET and
oxidative phosphorylation in mitochondria.
2. Dark Reactions
·
Use NADPH and ATP to synthesize CH2O
from H2O and CO2.
·
In eukaryotes,
occur in the stroma.
|
LIGHT
REACTIONS |
I. Photosynthetic
·
The primary
(photochemical) reactions of photosynthesis take place in RC. An RC from purple photosynthetic bacterium Rhodopseudomonas viridis is a transmembrane
protein containing a variety of chromophores. [bacteriochlorophyll
(Bchl) a and b, nonheme Fe(II),
bacteriopheophytin (BPheo)
b, ubiquinone, menaquinone.]
(BPheo is bacteriochlorophyll
in which the Mg2+ ion is replaced by
2 H+)
·
There are far
more chlorophyll molecules than are contained in RCs. These chlorophyll molecules do not
participate directly in the primary photochemical reaction but function as
antenna chlorophylls critical in capturing photons from sun light.
·
A
light-harvesting complex (LHC) (or antenna complex)
is a complex containing multiple membrane-bound hydrophobic proteins each
containing numerous chlorophylls and other pigments. The non-chlorophyll
pigments (i.e. accessory pigments such as carotenoids)
absorb at wavelengths where chlorophylls do not absorb strongly.
·
PRC is out-numbered by LHC.
·
Energy from
light captured by an antenna complex ® transfer from one to another antenna pigment molecule ® eventually trapped
by PRC (occurs in < 10-10 s, efficiency
> 90%) which has a lower energy excited state than that of antenna pigment
molecules. (Figs. 18-4, 7)
II. Two Photosystems and
Non-cyclic E Transport (Fig. 18-12)
·
All O2-evolving
photosynthetic cells contain photosystems I and II,
whereas non-oxygen-evolving photosynthetic bacteria contain only one system
(I).
·
Photosynthesis
requires the interaction of two light reactions, both of them can be driven by
light at <680 nm but only one (photosystem I) can
be driven by light at 680-700 nm.
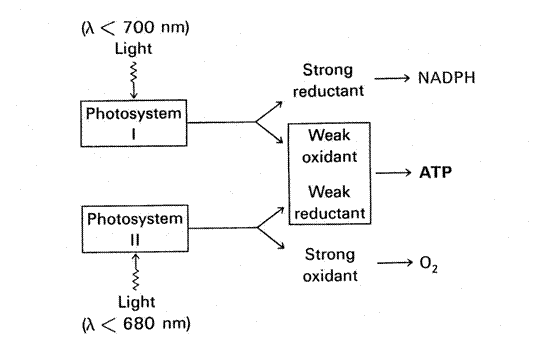
Taken from Stryer.
1. Photosystem I (formation of
NADPH)
·
Can be excited
at >680 nm but efficiency drops at >700 nm.
·
Absorbs 4
quanta, generates a strong reductant and a weak oxidant.
·
4 Χ [P700 + q →
P700* (= strong reductant) → P700+ (= weak
oxidant) + e]
·
4 Χ [P700+ + e (from P680*) → P700]
·
The strong
reductant provides 4 e to reduce 2 NADP+ to 2 NADPH.
·
NADPH is
formed by ferridoxin-NADP+ reductase in stroma.
·
The weak
oxidant receives 4 e from the weak reductant produced by photosystem
II (see below) to regenerate the ground state PSI P700.
2. Photosystem II (formation of O2 by oxygen-evolving center OEC)
·
Excitation
efficiency drops sharply at >680 nm.
·
Absorbs 4
quanta, generates a weak reductant and a strong oxidant.
·
4 Χ [P680 + q →
P680* (= weak reductant) → P680+ (= strong
oxidant) + e]
·
4 Χ [P680+ + e (from
water) → P680].
·
The strong
oxidant abstracts 4 e from 2H2O to generate 4H+
+ O2.
·
The weak
reductant provides 4 e to the weak oxidant formed by photosystem
I.
3. Formation of H+
Gradient and ATP (Figs. 18-11,
12)
·
The passage of
4e from the weak reductant of PSII to the weak
oxidant of PSI is coupled to the pumping of 8 H+
INTO thylakoid lumen.
In addition, the evolution of one O2 from 2H2O produces 4H+ in
the thylakoid lumen.
·
This passage
involves multiple redox-active factors, including chlorophylls, cytochromes, Q,
iron-sulfur clusters, and plastocyanin [a peripheral
membrane protein on the thylakoid luminal surface,
cycles between Cu(I) and Cu(II)].
·
This H+
gradient is driving the ATP synthesis by ATP synthase following a mechanism
similar to oxidative phosphorylation.
III.
Cyclic E Transport
(Fig. 18-12)
·
Involves only photosystem I.
·
Produces
ATP but not NADPH or O2.
·
Presumably
allow the cells to adjust the production of ATP relative to NADPH.
·
Regulation
of the partition between cyclic and noncyclic
pathways is not known.
EFFICIENCY OF ATP SYNTHESIS
Non-Cyclic E Transport
·
For every O2 formed (or 8
q absorbed):
·
4H+
are formed in thylakoid lumen from 2H2O
®
4H+ + O2.
·
In addition,
8H+ are pumped into thylakoid lumen when
4e pass from PSII to PSI.
·
So, 12/3 = 4
ATP can be formed from the H+ gradient resulting from the absorption
of 8 q.
·
Additionally,
the 2 NADPH formed by PSI are equivalent to another 6
ATP.
·
The overall
efficiency = 10 ATP per 8 q = 1.25 ATP/q.
Cyclic E Transport
For every 4 q
absorbed by PSI, 8 H+ are pumped into thylakoid lumen, resulting in the formation of (8/3) ATP.
Efficiency = (8/3)/4
= 0.67 ATP/q.